كتب مراقبة الرنا الرسول
اذا لم تجد ما تبحث عنه يمكنك استخدام كلمات أكثر دقة.
مراقبة الرنا الرسول (معلومة)
مراقبة جودة الرنا الرسول هي آليات تستخدمها الكائنات لضمان جودة ودقة جزيئات الرنا الرسول. توجد عدة آليات مراقبة في الخلية وتعمل هذه الآليات على عدة مراحل من حياة الرنا الرسول ويمكنها اكتشاف وتحديد جزيئات الرنا الرسول التي بها عيب في النسخ أو بها طفرة ثم تفكيكها.
ترجمة جزيئات الرنا الرسول المعيبة يمكن أن تؤدي إلى آثار ضارة على الكائن. تميل البروتينات غير الطبيعية الناتجة عن ترجمة جزيئات الرنا الرسول هذه إلى تكوين تكدساتٍ تؤدي إلى موت الخلية وإلى تكون الرقع النشوانية في الأنسجة.
نظرة عامة
ترجمة نسخ الرنا الرسول إلى بروتينات جزء حياتٌِّي في هدف علم الأحياء الجزيئي الرئيسي. مع ذلك، جزيئات الرنا الرسول عرضة للعديد من الأخطاء في دقة النسخ والتي تسبب بدورها أخطاءً في ترجمة الرنا إلى البروتين المطلوب. آليات مراقبة الرنا هي طرق تستخدمها الخلية لضمان جودة ودقة جزيئات الرنا الرسول. ويتم هذا عموما عبر وسم جزيئات الرنا الرسول المعيبة بطريقة محددة كي تتعرف عليها إنزيمات النوكليازات داخلية المنشأ وتقوم بتفكيكها.
وُثِّقت مراقبة الرنا الرسول لدى البكتيريا والخميرة. لدى حقيقيات النوى، تعمل هذه الآليات في كل من النواة والسيتوبلازم. ينتج عن فحوصات دقة جزيئات الرنا الرسول في النواة تفكيك النسخ المعالجة بشكل غير صحيح قبل تصديرها إلى السيتوبلازم. بعد ذلك تتعرض هذه النسخ إلى المزيد من المراقبة حين تصل السيتوبلازم. تفحص آليات المراقبة السيتوبلازمية نُسَخ الرنا بحثا عن تواجد أو غياب كودونات التوقف المبتسرة.
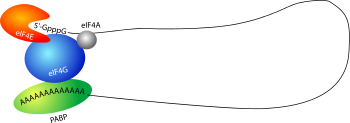
لدى حقيقيات النوى، توظيف الريبوسوم عند بداية الترجمة هو أول مستوى مراقبة جودة الرنا الرسول في السيتوبلازم. في سبيل توظيف الريبوسوم، من الضروري أن يكون الرنا الرسول على هيئة شبه دائرة ويكون ذيل عديد الأدينين 3" مرتبط بـPABP، وتكون القبعة 5" مرتبطة بعامل بدء الترجمة eIF4E، ثم يتآثر هاذين البروتينين مع عامل الترجمة eIF4G. هذه الآلية تضمن -قبل بدء الترجمة- أن الرنا الرسول سليم وكامل ويملك قبعة 5" وذيل عديد الأدينين 3"، ومنه جزيئات الرنا التي لا تحتوي على هاتين البنيتين لا تتم ترجمتها.
توجد ثلاث طرق مراقبة معروفة حاليا تعمل بها الخلايا وهي: تفكك الرنا الرسول المتوسَّط بلا معنى (NMD)، تفكك الرنا الرسول المتوسَّط بلا توقف (NSD)، وتفكك الرنا الرسولال المتوسَّط بلا مُضِي (NGD).
تفكك الرنا الرسول المتوسَّط بلا معنى
نظرة عامة
التفكك المتوسط بلا توقف (NSD) له دور في تحديد وتفكيك نسخ الرنا الرسول التي لا تملك كودون توقف. يمكن أن تنتج نسخ الرنا هذه من عدة آليات مثل التذييل بعديد الأدينيلات السابق لأوانه، إشارات التذييل بعديد الأدينين المشفَّرة داخل المنطقة المشفِّرة من الجين. انعدام كودون التوقف هو مشكل معتبر بالنسبة للخلايا، والريبوسومات المترجِمة للرنا الرسول تستمر في الترجمة وتقوم بترجمة ذيل عديد الأدينين ثم تعلق، وكنتيجة لذلك لا يستطيع الرنا الرسول الانفصال عنها. هذه الريبوسومات تصبح عالقة ومرتبطة بالرنا الرسول الذي لا يملك كودون توقف ولا تكون متاحة لترجمة جزئات رنا أخرى إلى بروتينات. تقوم آلية التفكك المتوسط بلا توقف بحل المشكلة عبر تخليص الريبوسومات العالقة ووسم ذلك الرنا الرسول للتفكيك بواسطة النوكليازات. يوجد مساران لآلية التفكك المتوسط بلا توقف وهذان المساران يعتمدان على وجود أو غياب البروتين Ski7.
مسار Ski7
يَنشط هذا المسار حين يكون بروتين Ski7 متوفرا في الخلية. يُعتقد أن البروتين Ski7 يرتبط بالموقع A الفارغ من الريبوسوم، ويسمح له هذا الارتباط بفصل وإخراج الرنا الرسول العالق الذي لا يحتوي كودون توقف مما يحرر الريبوسوم ويسمح له بترجمة نسخ أخرى. بعد الانفصال يكون بروتين Ski7 مرتبطا بالرنا الرسول وهذا الارتباط هو الإشارة التي تجعل الإكسوسوم في العصارة الخلوية يستهدف الرنا الرسول. يقوم مركب Ski7-إكسوسوم بإزالة ذيل عديد الأدينين بسرعة وهذا يسمح للإكسوسوم بتفكيك الرنا الرسول من الاتجاه 3" إلى 5".
مسار غير Ski7
لوحظ نوع ثانٍ من التفكك المتوسط بلا توقف لدى الخميرة. في هذا المسار، ينتج عن غياب بروتين Ski7 فقدانٌ لبروتينات PABP التي ترتبط بذيل عديد الأدينين بسبب الريبوسوم الذي يواصل الترجمة. إزالة بروتينات PABP يتسبب في فقدان القبعة 5"m7G التي تحمي النهاية 5"، وينتج عن هذا الفقدان تفكيك نسخة الرنا بواسطة النوكليازات الخارجية 5"-3" داخلية المنشأ مثل XrnI.
تفكك الرنا الرسول المتوسَّط بلا مضي
تفكك الرنا الرسول المتوسَّط بلا مضي (NGD) هو أحدث آليات المراقبة اكتشافا، ولهذا فالآلية غير مفهومة جيدا حاليا. مع كون الأهداف الحقيقية للتفكك المتوسَّط بلا مضي مفهومة بشكل ضعيف، يبدو أنها تتكون إلى حد كبير من جزيئات الرنا الرسول التي علقت الريبوسومات بها أثناء ترجمتها. يمكن أن يحدث هذا العلْق والتوقف بسبب عدة عوامل منها بنُى ثانوية قوية، والتي يمكن أن تسد وتمنع ماكينة الترجمة جسديا من التحرك على طول النسخة. يرتبط بروتينا Dom34/Hbs1 على الأرجح بالقرب من الموقع A للريبوسومات العالقة ويساهمان في تخليصها وإتاحتها لترجمات أخرى. في بعض الحالات، تُقص نسخة الرنا كذلك بالقرب من موقع العلق بطريقة النوكليازات الداخلية، لكن هوية النوكلياز الداخلي المسؤول عن ذلك مازالت محل نقاش. تُفكك جزيئات الرنا المقطَّع بشكل كامل بواسطة الإكسوسوم بطريقة 3" إلى 5" وبواسطة Xrn1 بطريقة 5" إلى 3".
حاليا طريقة تخليص الرنا الرسول من الريبوسومات مازلت غير معروفة، مع ذلك، البروتين Hbs1 ذو صلة قريبة من البروتين Ski7 الذي يلعب دورا واضحا في تخليص وتحرير الريبوسوم في التفكك المتوسط بلا توقف الذي يتم بواسطته. تم افتراض أن Hbs1 ربما يلعب دورا مماثلا في التفكك المتوسط بلا مضي.
تطور
من الممكن تحديد التاريخ التطوري لهذه الآليات عبر ملاحظة انحفاظ البروتينات المفتاحية التي تقوم بكل آلية. على سبيل المثال: Dom34/Hbs1 لهما دور في آلية بلا مضي، Ski7 له دور في آلية بلا توقف، و بروتينات eRF لها دور في آلية بلا معنى. لهذه الغاية تم القيام بأبحاث مكثفة بواسطة برنامج بلاست لتحديد تواجد هذه البروتينات في مختلف الأنواع من الكائنات. تم تحديد أن Hbs1 الخاص بآلية بلا مضي وeRF3 الخاص بآلية بلا معنى يتواجدان فقط لدى حقيقيات النوى. مع ذلك، Dom34 الخاص بآلية بلا مضي متواجد لدى حقيقيات النوى والعتائق، وهذا يوحي بأن آلية بلا مضي هي أول آليات مراقبة الرنا تطورا. يبدو أن Ski7 الخاص بآلية دون توقف محصور بشكل خاص لدى أجناس الخميرة وهذا يوحي بأن آلية بلا توقف هي أحدث آليات المراقبة تطورا، وهذا يترك آلية بلا معنى تلقائيا لتكون ثاني آليات المراقبة في التطور.






















")

















